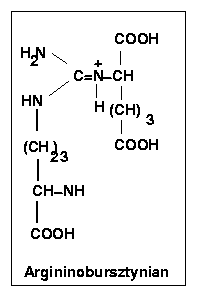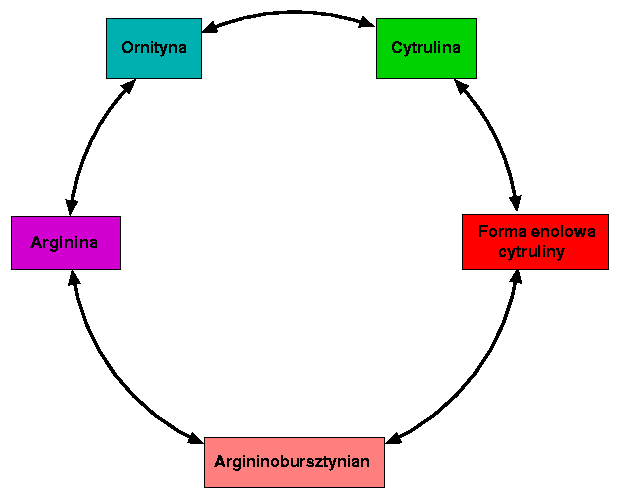
Przy udziale ATP i Mg2+ forma enolowa cyruliny uega kondensacji z asparaginianem, w wyniku czego tworzy się argininobursztynian. Przemianę tę katalizuje syntetaza arginobursztynianowa, która wymaga współdziałania ATP i Mg2+ ze wzgledu na endoergiczny charakter kondensacji. Reakcja jest łatwo odwracalna.
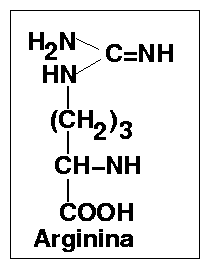
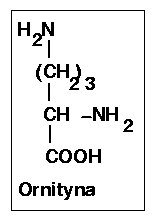
Reakcja ta jest kataliowana przez arginazę (ureohydrolazę argininową), enzym o absolutnej specyficzności w stosunku do L-argininy, wysokim optimum pH - 10,0 i wymagający obecności kofaktora w postaci jonów Mn2+ Enzym ten prowadzi do hydrolitycznego rozpadu argininy do mocznika ((NH2)2C=O) i ornityny i przez wytworzenie tej ostatniej zamyka obrót cyklu. Rekacja ta jest nieodwracalna, co przesądza o nieodwracalności cyklu.
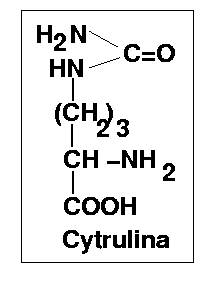
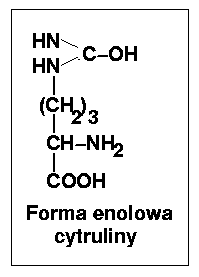
Cytrulina pozostaje w równowadze ze swoją formą enolową.
W reakcji tej zostaje przyłączony karbamoilofosforan (H2N-(C=O)-O~H2PO4), związek wysokiej energii, specyficzny dla tej endoergicznej przemiany i wytwarzany w przemianach towarzyszących cyklowi mocznikowemu z udziałem ATP. W przemianie tej służy jako dawca grupy karbamoilowej i energii. Enzymem katalizujacym tę reakcję jest karbamoilotransferaza ornitynowa, która nie wymaga dodatkowch kofaktorów i wykazuje znaczną specyficzność substratową. Reakcja ta jest odwracalna, a jej przebieg ze strony prawej na lewo prowadzi do tzw. fosforylacji substratowej.
Argininobursztynian ulega rozpadowi do argininy i fumaranu (trans HOOC-HC=CH-COOH ) pod wpływem działania enzymu liazy L - argininobursztynianowej. Jest to reakcja odwracalna, chociaż równowaga przesunięta na korzyść argininobursztynianu.